Bokutdrag fra boka Evolution in Four Dimensions
(Kapitlet: 'From Genes to Characters' -s61-65)
Bilde 1. Kolesterol ikke entydig begrep
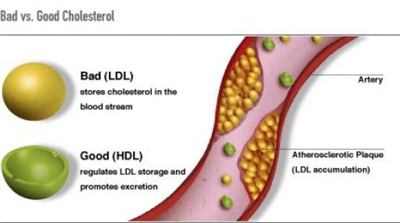 Tidligere ble det regnet med en enkel sammenheng: for høyt kolesterol ga forhøyet risiko for hjerte/kar-sykdommer. Nå har en funnet at sammenhengen ikke er så enkelt som før formodet: APOE-genet koder for et protein som hjelper å frakte fett rundt i blodet. Det har 3 vanlige alleler som kan kalles allele2, allele3 og allele 4. Undersøkelser indikerte at allele 4 kunne være 'dårlig' i forbindelse med hjerte/kar-sykdommer.
Tidligere ble det regnet med en enkel sammenheng: for høyt kolesterol ga forhøyet risiko for hjerte/kar-sykdommer. Nå har en funnet at sammenhengen ikke er så enkelt som før formodet: APOE-genet koder for et protein som hjelper å frakte fett rundt i blodet. Det har 3 vanlige alleler som kan kalles allele2, allele3 og allele 4. Undersøkelser indikerte at allele 4 kunne være 'dårlig' i forbindelse med hjerte/kar-sykdommer.
Om en betrakter kolesterol-nivå sammen med APOE-genotyper, blir bildet komplisert: a. Ikke alle med høyt kolesterol-nivå har økt risiko for hjerte/kar-sykdommer (om allele 3 er homo-zygote).b. Kombinasjon av de dårlige egenskapene høyt kolesterol og allele 4, er faktisk ikke det verste som kan forekomme. Det er de med høyt kolesterol og allele 2 som har størst risiko. c. Det er bare når du har middels eller lave kolesterol-verdier at det å ha allele 4, gir større risiko enn hos andre, med samme kolesterol-nivå. Derfor blir det for enkelt å hevde at høyt kolesterol-verdi og det å ha allele 4 i APOE-genet, isolert sett, fører til høyere risike for hjerte/kar-sykdommer. I tillegg kommer at andre gener kan påvirke risikoen -se under.
Det er flere forhold som vanskeliggjør muligheten for å forutsi fenotype fra vårt DNA. Det er at man noen ganger kan ha flere nettverk med samhandling fra ulike komponenter, som ender i å produsere identiske fenotyper. Visse aspekter av fenotypen synes forunderlig uavhengig av genetiske og miljømessige forskjeller. Så på ene siden kan man ha identiske gener som fører til meget ulike fenotyper, og på den andre side kan en ha ulike gener som produserer akkurat samme fenotype.
Bilde 2. Genregulerende nettverk (sjøpølse)
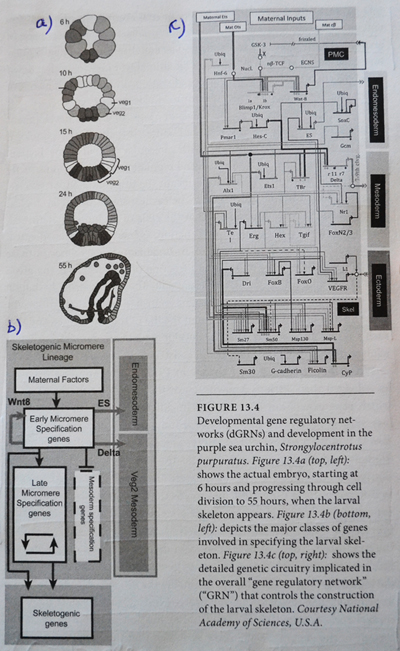
Dette er ikke nye oppdagelser (-selv om de ikke er så godt kjent-oversetters tilføyelse.) I 1940 og -50 åra presenterte den britiske ebryologen og genetikeren C. Waddington en visuell representasjon av denne ideen. En levende, nåværende illustrasjon av det intrikate og sofistikerte ved genetiske nettverk, ble tydelig da genetikere startet å benytte genetiske ingeniørkunster for å 'slå ut' et spesielt gen, og følge konsekvensene av denne utslagningen for utviklingen av embryoet. Man fant til sin overraskelse ut at å slå ut gener som hadde kjente effekter på viktige utviklingsveier, ofte ikke hadde noen effekt på den endelige fenotypen. På et vis kan genomet kompensere for fraværet av et gen.
Det er flere grunner til dette: i) Mange gener har duplikate kopier, så om begge alleler av en kopi blir slått ut, kan reserve-kopien kompensere. ii) Gener som normalt har andre funksjoner kan ta funksjonen til ett utslått gen. iii. Den dynamiske regulatoriske strukturen til underliggende nettverk, er slik at å slå ut enkelt-komponenter, blir uten effekt. Sluttproduktet blir likevel det samme, i det minste i de fleste omgivelser. 'Knock-out' eksperimenter viser at det er mye strukturell og funksjonell redundans i genomet. Samt at utviklingsstiene er så sterkt 'kanaliserte' at mange ulikheter i gener slår ut meget små forskjeller i fenotypen. Om et gen blir slått ut, kan strekket nettverket som forbinder dem fra andre gen-fester, etterlate landskapet temmelig uforandret.
Som konsekvens av dette: Siden vi nå vet mer om kompleksiteten som foregår mellom genet og fenotypen, er vi mindre forbauset over å finne nøytralitet av typen kommentert ovenfor. Og om det å slå ut et gen ikke skulle ha noen oppdagbar effekt, er det heller ingen grunn til at å endre en nukleotide her og der {-som ved punktmutasjoner, oversetters tilføyelse} skulle utgjøre noen forskjell. Det underliggende nettverk av samhandling som ligger under utvikling og vedlikehold av hvert trekk, er i stand til å tilpasse seg til eller kompensere for mange genetiske variasjoner. Det er derfor så mange potensielt ødeleggende effekter av stort antall variasjoner i DNA er kamuflert og nøytralisert.
Oversettelse og bilder ved Asbjørn E. Lund